Жаропонижающие средства для детей назначаются педиатром. Но бывают ситуации неотложной помощи при лихорадке, когда ребенку нужно дать лекарство немедленно. Тогда родители берут на себя ответственность и применяют жаропонижающие препараты. Что разрешено давать детям грудного возраста? Чем можно сбить температуру у детей постарше? Какие лекарства самые безопасные?
Основные закономерности наследственности, установленные для живых организмов, универсальны и в полной мере справедливы и для человека. Вместе с тем как объект генетических исследований человек имеет свои преимущества и недостатки.
Для людей невозможно планировать искусственные браки. Еще в 1923 г. Н.К. Кольцов отмечал, что "...мы не можем ставить опыты, мы не можем заставить Нежданову выйти замуж за Шаляпина только для того, чтобы посмотреть, какие у них будут дети". Однако эта трудность преодолима благодаря прицельной выборке из большого числа брачных пар тех, которые соответствуют целям данного генетического исследования.
В значительной мере затрудняет возможности генетического анализа человека большое число хромосом - 2п=4б. Однако разработка новейших методов работы с ДНК, метода гибридизации соматических клеток и некоторых других методов устраняют эту трудность.
Из-за небольшого числа потомков (во второй половине XX в. в большинстве семей рождалось по 2-3 ребенка) невозможен анализ расщепления в потомстве одной семьи. Однако в больших популяциях можно выбрать семьи с интересующими исследователя признаками. Кроме того, в некоторых семьях определенные признаки прослежены на протяжении многих поколений. В таких случаях возможен генетический анализ. Еще одна трудность связана с длительностью смены поколений у человека. Одно поколение у человека занимает в среднем 30 лет. И, следовательно, генетик не может наблюдать более одного-двух поколений.
Для человека характерен большой генотипический и фенотипический полиморфизм. Проявление многих признаков и болезней в сильной степени зависят от условий внешней среды. Следует отметить, что понятие "среда" для человека более широкое по сравнению с растениями и животными. Наряду с питанием, климатом и другими абиотическими и биотическими факторами, средой для человека являются и социальные факторы, трудно изменяемые по желанию исследователя. Вместе с тем, человека как генетический объект широко изучают врачи всех специальностей, что нередко помогает установить различные наследственные отклонения.
В настоящее время интерес и внимание к изучению генетики человека активно возрастают. Так, глобальная международная программа " Геном человека" имеет своей задачей изучение генома человека на молекулярном уровне. Для ее решения используются все новейшие современные методы генетики и медицины.
Какими же методами располагает генетика человека в настоящие дни? Их немало: генеалогический, близнецовый, цитогенетический, популяционно-статистический, биохимический, метод генетики соматических клеток и молекулярно-генетический. Рассмотрим подробнее каждый из них.
Относящийся к числу основных в генетике человека, этот метод опирается на генеалогию - учение о родословных. Его сутью является составление родословной и последующий ее анализ. Впервые такой подход был предложен английским ученым Ф. Гальтоном в 1865 г.
Генеалогический метод широко используется для решения как научных, так и прикладных проблем. Он позволяет выявить наследственный характер признака и определить тип наследования. Наряду с этим метод дает возможность установить сцепленное наследование, определить тип взаимодействия генов и пенетрантность аллелей. Генеалогический метод лежит в основе медико-генетического консультирования. Он включает два этапа: составление родословных и их генеалогический анализ.
Составление родословной. Сбор сведений о семье начинается с человека, называемого пробандом. Обычно это больной с изучаемым заболеванием. Дети одной родительской пары называются сибсами (братья-сестры). В большинстве случаев родословная собирается по одному или нескольким признакам. Родословная может быть полной или ограниченной. Чем больше поколений прослежено в родословной, тем она полнее и тем выше шансы на получение полностью достоверных сведений. Сбор генетической информации проводится путем опроса, анкетирования, личного обследования семьи. Опрос начинается обычно с родственников по материнской линии: бабушки и дедушки по материнской линии, с указанием внуков, детей каждого ребенка бабушки и дедушки. В родословную вносят сведения о выкидышах, абортах, мертворожденных, бесплодных браках и др.
При составлении родословной ведется краткая запись данных о каждом члене рода с указанием его родства по отношению к пробанду. Обычно указываются: фамилия, имя и отчество, дата рождения и смерти, возраст, национальность, место жительства семьи, профессия, наличие хронических заболеваний в семье, причину смерти умерших и др.
После сбора сведений составляют графическое изображение родословной, используя систему условных обозначений (рис. 2.1).
Выполняя эту работу, важно соблюдать следующие правила:
1. Составление родословной начинают с пробанда. Братья и сестры располагаются в порядке рождения слева направо, начиная со старшего.
2. Все члены родословной располагаются строго по поколениям в один ряд.
3. Поколения обозначаются римскими цифрами слева от родословной сверху вниз.
4. Арабскими цифрами нумеруется потомство одного поколения (один ряд) слева направо.
5. В связи с тем, что некоторые болезни проявляются в разные периоды жизни, указывается возраст членов семьи.
6. Отмечаются лично обследованные члены родословной.
Графическое изображение родословной может быть вертикально-горизонтальным или расположенным по кругу (в случае обширных данных). Схема родословной сопровождается описанием обозначений под рисунком, которое называется легендой (рис. 2.2).
Генетический анализ родословной
Задача генетического анализа - установление наследственного характера заболевания и типа наследования, выявление гетерозиготных носителей мутантного гена, а так же прогнозирование рождения больных детей в семьях с наследственной патологией.
Анализ родословной включает следующие этапы: 1. Установление, является ли данный признак или заболевание единичным в семье или имеется несколько случаев (семейный характер). Если признак встречается несколько раз в разных поколениях, то можно предполагать, что этот признак имеет наследственную природу. 2. Определение типа наследования признака. Для этого анализируют родословную, учитывая следующие моменты:
1) встречается ли изучаемый признак во всех поколениях и многие ли члены родословной обладают им;
2) одинакова ли его частота у лиц обоих полов и у лиц какого пола он встречается чаще;
3) лицам какого пола передается признак от больного отца и больной матери;
4) есть ли в родословной семьи, в которых у обоих здоровых родителей рождались больные дети, или у обоих больных родителей рождались здоровые дети;
5) какая часть потомства имеет наследуемый признак в семьях, где болен один из родителей.
Аутосомно-доминантное наследование характеризуется тем, что мутантный ген связан с аутосомой и проявляется как в гомозиготном (АЛ), так и в гетерозиготном (Аа) состояниях. В силу этого прослеживаются следующие особенности наследования:
1) передача патологии от больных родителей к детям;
2) оба пола поражаются в равных пропорциях;
3) здоровые члены семьи обычно имеют здоровое потомство;
4) отец и мать одинаково передают мутантный ген дочерям и сыновьям. Возможна передача болезни от отца к сыну.
Клинические проявления болезни могут значительно варьировать в зависимости от экспрессивности и пенетрантности гена. Экспрессивностью называется степень выраженности гена (в нашем случае - тяжесть заболевания). При высокой экспрессивности гена развивается тяжелая, часто с летальным исходом форма заболевания, при низкой - внешне человек здоров. Под пенетрантностю понимается частота проявления мутантного гена среди его носителей. Она определяется отношением числа особей, имеющих данную болезнь (или признак), к числу особей, имеющих данный ген, выраженным в процентах. Например, пенетрантность атеросклероза 40%, синдрома Марфана 30%, ретинобластомы 80% и т.д.
В зависимости от типа наследования общая картина родословной выглядит по-разному
При аутосомно-рецессивиом типе наследования мутанный ген проявляет свое действие только в гомозиготном состоянии По это л причине в гетерозиготном состоянии он может существовать во многих поколениях, не проявляясь фенотипически.
При данном типе наследования заболевание встречаете в родословной редко и не во всех поколениях. Вероятность заболевания у девочек и мальчиков одинакова. Признак может проявиться у детей, родит ели которых были здоровы, но являлось гетерозиготными носителями мутантного гена. Возможны несколько вариантов таких браков:
1) мать аа х отец аа - у таких родителей все дети будут больными (аа);
2) мать Аа х отец аа - 50 % детей будут больными (генотип аа) и 50 % фенотипически здоровыми (генотип Аа), но будут являться гетерозиготными носителями дефектного гена;
3) мать Аа х отец Аа - 25 % детей будут больными (генотип аа), 75 % фенотипически здоровыми (генотипы АА и Аа), но 50 % из них будут носителями мутантного гена (генотип Аа).
Экспрессивность и пенетрантность колеблются в широких пределах (от О до 100%) и сильно зависят от условий внешней среды. По аутосомно-доминантному типу наследуется полидактилия (шестипалость), брахидактилия (короткопалость), ахондроплазия (карликовость), синдром Марфана ("паучьи пальцы") и другие заболевания (рис. 2.3).
При доминантном типе наследования, если один из родителей болен (Аа), вероятность рождения больного ребенка равна 50% при условии полной пенетрантности гена. В случае гетерозиготности обоих родителей (Аа х Аа) больные дети могут родиться с вероятностью 75%. Многие аутосомно-доминантные заболевания в гомозиготном состоянии протекают в более тяжелой форме, чем у гетерозигот. Однако в практике нередки случаи, когда носители доминантного гена остаются фенотипически здоровыми. В результате вид родословной изменяется и появляются пропуски поколений.
Носительство доминантного гена без фенотипического проявления можно заподозрить у одного из родителей, если среди его потомков появились больные с такой же доминантной патологией. Когда у здоровых родителей появился больной ребенок и в родословной есть другие случаи этой болезни, то правомочно предполагать, что у одного из родителей больного был дефектный ген, который не пенетрировал, но был передан потомку.
Доминантный ген может обладать разной степенью экспрессивности, что затрудняет установление аутосомно-доминантного типа наследования. Рассмотрим это на примере наследственной патологии соединительной ткани - синдрома Марфана.
Известно, что частота возникновения наследственных рецессивно-аутосомных болезней находится в прямой зависимости от степени распространенности мутантного гена среди населения. Частота таких болезней особенно повышается в изолятах и среди населения с высоким процентом кровнородственных браков. Такие браки отрицательно влияют на потомство, на это указывает тот факт, что умственная отсталость среди детей от родственных браков в 4 раза выше, чем в семьях с неродственными браками.
При аутосомно-рецессивном типе наследования (как и при аутосомно-доминантном) возможны различная степень экспрессивности и пенетрантности признака. К заболеваниям с аутосомно-рецессивным типом наследования относятся многие болезни обмена веществ, среди которых фенилкетонурия, галактоземия, альбинизм (рис.2.4), муковисцидоз и др. Установлено, что рецессивные заболевания чаще диагностируются в раннем возрасте.
Наследование заболеваний, сцепленных с полом, определяется тем, что мутантный ген расположен в X или Y-хромосоме. Известно, что у женщин имеются две Х-половые хромосомы, а у мужчин - одна Х- и одна Y-хромосома. У человека в Х-хромосоме локализовано более 200 генов. Гены, расположенные в хромосоме X, могут быть рецессивными или доминантными.
У женщин мутантный ген может находиться в обеих Х-хромосомах или только в одной из них; в первом случае она гомозиготна, во втором - гетерозиготна. Мужчины, являясь гемизиготными (имеют только одну хромосому X), передают ее только дочерям и никогда - сыновьям. Любой ген, как доминантный, так и рецессивный, локализованый в его Х-хромосоме, обязательно будет проявляться. В этом главная особенность Х-сцепленного наследования.
Для Х-сцепленного рецессивного наследования характерны следующие особенности:
1) заболевание встречается чаще у лиц мужского пола;
2) от здоровых родителей могут родиться больные дети (если мать гетерозиготна по мутантному гену);
3) больные мужчины не передают заболевание своим сыновьям, но их дочери становятся гетерозиготными носителями болезни;
4) больные женщины могут родиться только в семьях, где отец болен, а мать гетерозиготна по мутантному гену.
Рассмотрим несколько примеров, когда в Х-хромосоме локализован рецессивный ген. Если в брак вступает здоровая женщина и больной мужчина, то в такой семье все дети будут здоровыми, а дочери получат от отца одну Х-хромосому с мутантным геном и будут гетерозиготными носителями (т.к. вторую нормальную Х-хромосому они получат от матери). В том случае, если в брак вступают здоровый мужчина и женщина - носительница патологического гена, то вероятность рождения больного мальчика составит 50% от всех мальчиков и 25% - от всех детей.
Вероятность рождения больных девочек очень низка и возможна лишь, если отец болен, а мать гетерозиготна по мутантному гену. В такой семье половина мальчиков будут больными. Среди девочек болезнь проявится у половины, а другая половина будет нести дефектный ген.
Классическим примером рецессивного, сцепленного с полом наследования, может служить гемофилия. Больные страдают повышенной кровоточивостью. Причина - недостаточное содержание в крови факторов свертывания крови. На рис. 2.5 изображена родословная семьи с гемофилией
Анализ родословной показывает, что болеют только мальчики. (II - 1,4; III - 7,15). Отсюда можно предположить, что ген гемофилии сцеплен с полом. Больные дети чаще рождаются от здоровых родителей и, следовательно, ген болезни рецессивный.
Известно, что гемофилия широко распространена в королевских семьях Европы. Это связано с заключением близкородственных браков. В результате возникшие мутации сохранялись внутри семьи. Королева Виктория в Англии была носительницей гена гемофилии. Ее сын Леопольд родился гемофиликом. Через своих дочерей и внуков королева Виктория передала ген гемофилии Вольдемару и Генриху Прусским, Фридриху Гессенскому, царевичу Алексею Романову, Рупрехту Тех-Атлонскому, двум баттенбергским и двум испанским принцам (рис 2.6). Кроме гемофилии, в Х-хромосоме локализованы рецессивные гены, обусловливающие миопатию Дюшенна, некоторые формы дальтонизма и др. заболевания.
При локализации в хромосоме X доминантного гена тип наследования называется Х-сцепленным доминантным. Для него характерны следующие признаки:
1) болеют как мужчины, так и женщины, однако больных женщин в два раза больше, чем больных мужчин;
2) заболевание прослеживается в каждом поколении;
3) если болен отец, то все его дочери будут больными, а все сыновья здоровыми;
4) если мать больна, то вероятность рождения больного ребенка равна 50 % независимо от пола;
5) больными дети будут только тогда, если болен один из родителей;
6) у здоровых родителей все дети будут здоровы.
По Х-сцеплснному доминантному типу наследуются фосфатемия (недостаток фосфата в крови), коричневая окраска эмали зубов и др.
Имеет свои особенности и Y-сцепленное наследование.
В Y-хромосоме у мужчин локализовано немного генов. Они передаются только сыновьям и никогда дочерям (голандрическое наследование). С Y-хромосомой у мужчин наследуются такие признаки, как гипертрихоз (наличие волос по краю ушных раковин), кожные перепонки между пальцами ног, развитие семенников, интенсивность роста тела, конечностей и зубов. Характерные особенности наследования с Y-хромосомой можно видеть на рис. 2.7.
В основе этого метода лежит составление и анализ родословных. Этот метод широко применяют с древних времен и до наших дней в коневодстве, селекции ценных линий крупного рогатого скота и свиней, при получении чистопородных собак, а также при выведении новых пород пушных животных. Родословные человека составлялись на протяжении многих столетий в отношении царствующих семейств в Европе и Азии.
Как метод изучения генетики человека генеалогический метод стали применять только с начала XX столетия, когда выяснилось, что анализ родословных, в которых прослеживается передача из поколения в поколение какого-то признака (заболевания), может заменить собой фактически неприменимый в отношении человека гибридологический метод.
При составлении родословных исходным является человек - пробанд, родословную которого изучают. Обычно это или больной, или носитель определенного признака, наследование которого необходимо изучить. При составлении родословных таблиц используют условные обозначения, предложенные Г. Юстом в 1931 г. (рис. 6.24). Поколения обозначают римскими цифрами, индивидов в данном поколении - арабскими.
Рис. 6.24. Условные обозначения при составлении родословных (по Г. Юсту)
С помощью генеалогического метода может быть установлена наследственная обусловленность изучаемого признака, а также тип его наследования (аутосомно-доминантный, аутосомно-рецессивный, X-сцепленный доминантный или рецессивный, Y-сцепленный). При анализе родословных по нескольким признакам может быть выявлен сцепленный характер их наследования, что используют при составлении хромосомных карт. Этот метод позволяет изучать интенсивность мутационного процесса, оценить экспрессивность и пенетрантность аллеля. Он широко используется в медико-генетическом консультировании для прогнозирования потомства. Однако необходимо отметить, что генеалогический анализ существенно осложняется при малодетности семей.
Родословные при аутосомно-доминантном наследовании. Для аутосомного типа наследования в целом характерна равная вероятность встречаемости данного признака как у мужчин, так и у женщин. Это обусловлено одинаковой двойной дозой генов, расположенных в аутосомах у всех представителей вида и получаемых от обоих родителей, и зависимостью развивающегося признака от характера взаимодействия аллельных генов.
При доминировании признака в потомстве родительской пары, где хотя бы один родитель является его носителем, он проявляется с большей или меньшей вероятностью в зависимости от генетической конституции родителей (рис. 6.25).

Рис. 6.25. Вероятность появления потомков с доминантным признаком от различных супружеских пар (/-III )
Если анализируется признак, не влияющий на жизнеспособность организма, то носители доминантного признака могут быть как гомо-, так и гетерозиготами. В случае доминантного наследования какого-то патологического признака (заболевания) гомозиготы, как правило, нежизнеспособны, а носители этого признака - гетерозиготы.
Таким образом, при аутосомно-доминантном наследовании признак может встречаться в равной мере у мужчин и у женщин и прослеживается при достаточном по численности потомстве в каждом поколении по вертикали. Анализируя родословные, необходимо помнить о возможности неполного пенетрирования доминантного аллеля, обусловленной взаимодействием генов или факторами среды. Показатель пенетрантности может быть вычислен как отношение фактического числа носителей признака к числу ожидаемых носителей этого признака в данной семье. Необходимо также помнить, что некоторые заболевания проявляются не сразу с момента рождения ребенка. Многие болезни, наследуемые по доминантному типу, развиваются лишь в определенном возрасте. Так, хорея Гентингтона клинически проявляется к 35-40 годам, поздно проявляется и поликистоз почек. Поэтому при прогнозировании подобных заболеваний в расчет не принимаются братья и сестры, не достигшие критического возраста.
Первое описание родословной с аутосомно-доминантным типом наследования аномалии у человека было дано в 1905 г. В ней прослеживается передача в ряду поколений брахидактилии (короткопалости). На рис. 6.26 приведена родословная с этой аномалией. На рис. 6.27 изображена родословная с ретинобластомой в случае неполной пенетрантности.
Родословные при аутосомно-рецессивном наследовании. Рецессивные признаки проявляются фенотипически лишь у гомозигот по рецессивным аллелям. Эти признаки, как правило, обнаруживаются у потомков фенотипически нормальных родителей - носителей рецессивных аллелей. Вероятность появления рецессивного потомства в этом случае равна 25%. Если один из родителей имеет рецессивный признак, то вероятность проявления его в потомстве будет зависеть от генотипа другого родителя. У рецессивных родителей все потомство унаследует соответствующий рецессивный признак (рис. 6.28).

Рис. 6.26. Родословная (А ) при аутосомно-доминантном типе наследования (брахидактилия - Б )
Для родословных при аутосомно-рецессивном типе наследования характерно, что признак проявляется далеко не в каждом поколении. Чаще всего рецессивное потомство появляется у родителей с доминантным признаком, причем вероятность появления такого потомства возрастает в близкородственных браках, где оба родителя могут являться носителями одного и того же рецессивного аллеля, полученного от общего предка. Примером аутосомно-рецессивного наследования является родословная семьи с псевдогипертрофической прогрессивной миопатией, в которой часты близкородственные браки (рис. 6.29). Обращает внимание распространение заболевания в последнем поколении по горизонтали.

Рис. 6.27. Родословная с ретинобластомой в случае неполной пенетрантности

Рис. 6.28. Вероятность появления потомков с рецессивным признаком
от различных супружеских пар (I-IV )
Родословные при доминантном Х-сцепленном наследовании признака. Гены, расположенные в Х-хромосоме и не имеющие аллелей в Y-хромосоме, представлены в генотипах мужчин и женщин в разных дозах. Женщина получает две свои Х-хромосомы и соответствующие гены как от отца, так и от матери, а мужчина наследует свою единственную Х-хромосому только от матери. Развитие соответствующего признака у мужчин определяется единственным аллелем, присутствующим в его генотипе, а у женщин он является результатом взаимодействия двух аллельных генов. В связи с этим признаки, наследуемые по Х-сцепленному типу, встречаются в популяции с разной вероятностью у мужского и женского пола.
При доминантном Х-сцепленном наследовании признак чаще встречается у женщин в связи с большей возможностью получения ими соответствующего аллеля либо от отца, либо от матери. Мужчины могут наследовать этот признак только от матери. Женщины с доминантным признаком передают его в равной степени дочерям и сыновьям, а мужчины - только дочерям. Сыновья никогда не наследуют от отцов доминантного Х-сцепленного признака.

Рис. 6.29. Родословная при аутосомно-рецессивном типе наследования (псевдогипертрофическая прогрессирующая миопатия)
Примером такого типа наследования служит описанная в 1925 г. родословная с фолликулярным кератозом - кожным заболеванием, сопровождающимся потерей ресниц, бровей, волос на голове (рис. 6.30). Характерным является более тяжелое течение заболевания у гемизиготных мужчин, чем у женщин, которые чаще всего являются гетерозиготами.
При некоторых заболеваниях наблюдается гибель мужчин-гемизигот на ранних стадиях онтогенеза. Тогда в родословных среди пораженных должны быть только женщины, в потомстве которых отношение пораженных дочерей, здоровых дочерей и здоровых сыновей равно 1:1:1. Мужские доминантные гемизиготы, не погибающие на очень ранних стадиях развития, обнаруживаются в самопроизвольных абортах или среди мертворожденных. Такими особенностями наследования у человека характеризуется пигментный дерматоз.
Родословные при рецессивном Х-сцепленном наследовании признаков. Характерной особенностью родословных при данном типе наследования является преимущественное проявление признака у гемизиготных мужчин, которые наследуют его от матерей с доминантным фенотипом, являющихся носительницами рецессивного аллеля. Как правило, признак наследуется мужчинами через поколение от деда по материнской линии к внуку. У женщин он проявляется лишь в гомозиготном состоянии, вероятность чего возрастает при близкородственных браках.
Наиболее известным примером рецессивного Х-сцепленного наследования является гемофилия. Наследование гемофилии типа А представлено в родословной потомков английской королевы Виктории (рис. 6.31).

Рис. 6.30. Родословная при Х-сцепленном доминантном типе наследования (фолликулярный кератоз)

Рис. 6.31. Родословная при Х-сцепленном рецессивном типе наследования (гемофилия типа А)
Другим примером наследования по данному типу является дальтонизм - определенная форма нарушения цветоощущения.
Родословные при Y-сцепленном наследовании. Наличие Y-хромосомы только у представителей мужского пола объясняет особенности Y-сцепленного, или голандриче-ского, наследования признака, который обнаруживается лишь у мужчин и передается по мужской линии из поколения в поколение от отца к сыну.

Рис. 6.32. Родословная при Y-сцепленном (голандрическом) типе наследования
Одним из признаков, Y-сцепленное наследование которого у человека все еще обсуждается, является гипертрихоз ушной раковины, или наличие волос на внешнем крае ушной раковины. Предполагают, что в коротком плече Y-хромосомы кроме этого гена находятся гены, определяющие мужской пол. В 1955 г. у мыши описан определяемый Y-хромосомой трансплантационный антиген, названный HY. Возможно, он является одним из факторов половой дифференцировки мужских гонад, клетки которых имеют рецепторы, связывающие этот антиген. Связанный с рецептором антиген активизирует развитие гонады по мужскому типу (см. разд. 3.6.5.2; 6.1.2). Этот антиген в процессе эволюции остался почти неизменным и встречается в организме многих видов животных, в том числе и человека. Таким образом, наследование способности к развитию гонад по мужскому типу определяется голандрическим геном, расположенным в Y-хромосоме (рис. 6.32).
У человека как объекта генетического исследования почти нет никаких преимуществ перед другими объектами.
Напротив, много препятствий, затрудняющих изучение его генетики: 1) невозможность произвольного скрещивания в эксперименте; 2) позднее наступление половой зрелости; 3) малое число потомков в каждой семье; 4) невозможность уравнивать условия жизни для потомства; 5) отсутствие точной регистрации проявления наследственных свойств в семьях и отсутствие гомозиготных линий; 6) большое число хромосом; 7) и самым главным затруднением изучения генетики человека в капиталистическом обществе является социальное неравенство, что затрудняет реализацию наследственных потенций человека.
Несмотря на указанные затруднения, генетика разработала некоторые методы, которые позволяют шаг за шагом изучать наследственность и наследование у человека. Существует несколько методов исследования: генеалогический, цитогенетический, близнецовый, онтогенетический и популяционный.
Следует иметь в виду, что любой признак, независимо от того, является ли он признаком дикого типа, т. е. относится к норме, или связан с каким-либо заболеванием, может служить моделью для изучения наследственности. Оградить человека от наследственных болезней или поражения его наследственности так же важно, как и выяснить наследование нормы. В настоящее время генетические методы разработаны главным образом в отношении морфологических признаков, которые генетически определяются достаточно четко (брахидактилия, альбинизм, дальтонизм, пятнистость кожи и волос и т. д.).
Генетическое исследование психических свойств все еще остается проблематичным, так как для них не найдены элементарные критерии признака в генетическом смысле. Почти все признаки Психической и творческой деятельности человека настолько комплексны и сложны, а также в сильной степени обусловлены внешними, в том числе и социальными, факторами, что генетический анализ этих свойств пока трудно осуществим, хотя наследственная их обусловленность не вызывает сомнения.
Можно сказать, что значительное большинство признаков, характеризующих вид Homo sapiens, может изучаться как количественные и сложные физиологические признаки, т. е. признаки, не проявляющие дискретного характера в онтогенезе. Эти признаки контролируются системой генотипа (полигенно). И пока эта система не разгадана хотя бы на примере просто организованных организмов, проблема признаков поведения остается малодоступной для генетического анализа. Напротив, мутантные признаки, выходящие за границы характеристики видовых признаков, служат хорошими генетическими моделями изучения наследственности и наследования в норме.
На дискретные мутантные признаки нельзя смотреть как на признаки только патологические, якобы не имеющие приспособительного значения. Возможно, что само появление человека с развитыми полушариями коры головного мозга, вертикальным положением тела, дискретной речевой сигнализацией является следствием крупных мутаций. В пользу этого свидетельствует очень
короткий промежуток времени эволюции человека, за который мелкие мутации вряд ли могли накопиться в таком количестве и дать такой значительный эволюционный эффект. Разумный человек для природы столь же «необычен», как домашняя курица, несущая 365 яиц в год вместо 10-15, или рекордистка-корова, дающая 16 тыс. кг молока в год вместо 600-700 кг.
Разделение признаков на нормальные и мутантные применительно к человеку и животным необходимо для познания эволюции человека и патологических явлений.
Совокупность видовых признаков человека и животных определяется системой генотипа, сложившейся под влиянием всех факторов отбора в процессе эволюции. Мутации, пребывающие в гетерозиготном состоянии у человека, по-видимому, так же необходимы, как и у животных, для поддержания их в популяции.
Самым опасным в разработке научных методов исследования животных и человека, особенно его способностей, является антропоморфический момент, т. е. выдача желаемого за действительность.
Генеалогический метод
Анализ наследования человека на основе составления родословной - генеалогии был предложен Ф. Гальтоном.
Генеалогический метод представляет собой изучение наследования свойств человека по родословным (педигри). Данный метод применим, если известны прямые родственники - предки обладателя наследственного признака (пробанда) по материнской и отцовской линиям в ряду поколений и имеется достаточное число потомков в каждом поколении, или в том случае, когда имеются данные по достаточному числу разных семей, позволяющему выявить сходство родословных. Данные по совокупности сходных родословных подвергают статистической обработке.
Получившая наибольшее распространение система обозначения родословных человека была предложена Г. Юстом в 1931 г.
На основе большого числа проанализированных семей составляют родословные и производят математические расчеты соответственно типу наследования того или иного признака - доминантному или рецессивному, часто и не часто встречающейся мутации, сцепленному или не сцепленному с полом и т. д. Здесь мы не будем касаться приложения математического метода к данному анализу, отметим только, что весь этот формальный анализ основан на элементарных генетических закономерностях наследования.
Схемы родословной наследования дохминантного аутосомного гена, определяющего какой-либо признак, например заболевание (хондродистрофическая карликовость, буллезный эпидермолиз - свойство кожи образовывать большие пузыри при небольших травмах, ретинобластома и т. д.), или морфологический недостаток, например короткопалость (брахидактилия - отсутствие двух дистальных фаланг в пальцах).

Наследование признаков, определяемых рецессивными генами (рецессивное наследование), анализируется несколько сложнее, при составлении схем родословных.
Например, двоих в семье, появление двоих больных детей равно произведению вероятностей, т. е. 0,25 X 0,25, т. е. 6,25%.
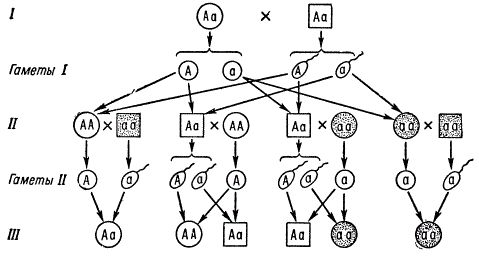
Часто встречающиеся рецессивные аутосомные гены при условии, если носители их (аа) способны вступать в брак и давать потомство, будут находиться в высокой концентрации в популяции. В таком случае становятся очень вероятными браки аа X Аа, в потомстве от которых наследование данного признака будет имитировать наследование по доминантному типу 1:1. Однако, зная тип наследования и проявления тех и других генов даже в случае малочисленных семей, но при достаточном числе таких семей, можно установить истинный характер наследования.
Наследование генов, полностью сцепленных с полом, т. е. находящихся в негомологичных сегментах, и частично сцепленных с полом - локализованных в гомологичных сегментах X- и Y-xpoмосом, подчиняется установленным для половых хромосом закономерностям. Для доминантных и рецессивных генов это наследование будет определяться по-разному в зависимости от того, где данный ген локализован - в гомологичном или негомологичном сегменте X- и Y-хромосомы и каким образом он передается. Так, доминантный ген, вызывающий перепончатость пальцев, находящийся в негомологичном сегменте Y-хромосомы, наследуется от отцов и проявляется только у мужчин.
Для частично сцепленных с полом доминантных генов, находящихся в гомологичных сегментах половых хромосом, анализ несколько более затруднен, но также возможен. Примером сцепленного с полом наследования рецессивного признака является наследование гемофилии. В передаче этого признака в поколениях имеется прерывность; пораженные мужчины являются потомками здоровых матерей, которые были гетерозиготами по данному гену; больные гемофилией женщины могут быть потомками больного отца и больной или здоровой матери.

У человека найдено около 50 сцепленных с полом рецессивных генов. Интересно, что около половины из них обусловливают заболевание глаз. Уже с давних времен было известно, что степень передачи наследственных признаков в родственных (инбридинг) и неродственных браках (аутбридинг) различна. После того, как генетика установила закономерности более частого проявления рецессивных генов при инбридинге, нет необходимости пространно доказывать вред родственных браков. Чем выше коэффициент инбридинга, тем больше вероятность появления наследственных болезней в поколениях. В разных странах среди разных народов и классов общества, а также в разные эпохи родственные браки (между двоюродными, троюродными братьями и сестрами) встречаются с разной частотой. Так, например, в деревнях на островах Фиджи количество родственных браков достигает 29,7%, в некоторых кастах Индии - 12,9, в Японии (Нагасаки) - 5,03, в Голландии - 0,13-0,159, в Португалии- 1,40, в США (Балтимора) - 0,05%, и т. д. Процент родственных браков колеблется в отдельных районах одной и той же страны в зависимости от уклада жизни.
Вредность родственных браков мало заметна в отдельных родословных, но при сравнительном статистическом анализе болезней и смертностей она выступает с полной очевидностью.

Яркий пример выявления рецессивного гена при родственном браке.

В этой родословной родство поддерживается через бракосочетание сибсов (братья - сестры) разной степени родства. От двух родственных браков (четвероюродные сибсы) появилось в одной семье 4 ребенка из 8, а в другой - 2 из 5, страдающих наследственной амавротической идиотией. К. Штерн предполагает, что один из двух общих предков этих линий передал данный рецессивный ген через три поколения каждому из четырех родителей.
Иногда заболевание и смертность детей от родственных браков превышают на 20-30% таковые от неродственных браков. Очевидно, что причина рассматриваемого явления генетическая, а именно: большая вероятность проявления наследственных заболеваний и смертности в результате гомозиготизации рецессивных генов, определяющих физиологические недостаточности и смертность (летальные и полулетальные гены).
Итак, генеалогический метод является весьма ценным методом, однако его значение в исследованиях тем больше, чем точнее и глубже составлены родословные. По мере роста цивилизации и более точной регистрации родословных роль этого метода в генетике человека будет возрастать.
Близнецовый метод
Близнецами называют потомство, состоящее из одновременно родившихся особей у одноплодных животных (человек, лошадь, крупный рогатый скот, овцы и др.).
Близнецы могут быть однояйцевыми и разнояйцевыми.
Идентичные, или однояйцевые, близнецы (ОБ) развиваются из одного яйца, оплодотворенного одним сперматозоидом, когда из зиготы вместо одного зародыша возникают два или более (полиэмбриония). В силу того, что митотическое деление зиготы дает два равнонаследственных бластомера, однояйцевые близнецы, сколько бы их ни развивалось, должны быть наследственно идентичны и одного пола. Это явление представляет собой пример бесполого, а точнее, вегетативного размножения животных.
Разнояйцевые близнецы (РБ) развиваются из одновременно овулировавших разных яйцеклеток, оплодотворенных разными сперматозоидами. И так как разные яйцеклетки и сперматозоиды могут нести различные комбинации генов, то разнояйцевые близнецы могут быть наследственно столь же разными, как и дети одной и той же супружеской пары, родившиеся в разное время. Разнояйцевые близнецы могут быть одного (РБо) или разного пола (РБр).
Чаще в литературе вместо термина «разнояйцевые близнецы» (РБ) употребляют термин «двуяйцевые близнецы» (ДБ), так как двойни встречаются чаще. Однако термин «разнояйцевые близнецы» лучше подчеркивает разницу между ОБ и РБ; однояйцевые близнецы также чаще рождаются двойнями.
Судя по данным, полученным на млекопитающих, для объяснения образования ОБ у человека может быть несколько гипотез:
- расхождение бластомеров при первом дроблении зиготы и раздельное развитие зародыша из этих бластомеров;
- разделение группы клеток на стадии бластоциста (до гаструляции);
- разделение зародышей на ранней стадии гаструляции. Наиболее вероятным путем предполагают второй.

Число близнецов в одних родах у человека колеблется: чаще всего встречаются двойни, реже тройни, еще реже - четверни, совсем редко - пятерни. По данным И. И. Канаева, за последние 150 лет в США установлено четыре случая родов пятерни, в Канаде - два случая. Факт рождения ОБ - пятерни девочек, доживших до взрослого состояния, - известен в семье канадского фермера Дионн (1934 г.). Рассчитано, что пятерни рождаются один раз на 54 700 816 родов, шестерни - на 4712 млн. родов, семерни известны только как исключение. В среднем частота рождения близнецов составляет 1% с колебаниями в пределах 0,5-1,5%. Близнецы менее жизнеспособные, и поэтому их количество при рождении меньше, чем при зачатии, а во взрослом состоянии меньше, чем при рождении.
Расчет частоты ОБ по отношению к РБ делается исходя из теоретического соотношения однополых и разнополых пар РБ при рождении близнецов: 25%♀♀ + 50%♀♂ + 25%♂♂ вычитание числа пар разного пола из общего числа всех пар одинакового пола (мужского и женского) даст разницу, составляющую число пар ОБ, которая в среднем колеблется от 21 до 33,4% всех близнецов.
Для использования близнецов в генетических исследованиях очень важно точно диагностировать тип ОБ и тип РБ. Диагностика производится на основании следующих критериев:
- ОБ обязательно одного пола, РБ могут быть как одного пола, так и разных полов;
- ОБ имеют, как правило, один общий хорион, РБ - разные хорионы;
- реципрокная трансплантация тканей у ОБ столь же успешна, как и автотрансплантация, у РБ она невозможна;
- наличие сходства (конкордантности) у ОБ и несходства (дискордантности) у РБ по многим признакам.
Для диагностики следует выбирать признаки, четко наследующиеся и менее всего подверженные изменению под влиянием факторов среды; к таким признакам относятся группы крови, пигментация глаз, кожи и волос, кожный рельеф (отпечатки кончиков пальцев, ладоней, ступней и др.). Если по одному-двум таким признакам выявлено различие близнецов, то они, как правило, являются РБ.
Все сомнительные случаи диагностики близнецов могут быть вызваны либо нарушением развития одного из партнеров ОБ, либо сходством родителей по ряду признаков. Однако последнее встречается чрезвычайно редко. Следует заметить, что нарушение развития одного из партнеров ОБ обычно объясняют неодинаковым действием факторов внутриутробной жизни и возникновением соматических мутаций на ранних стадиях эмбрионального развития, до закладки органов. Различного рода генные и хромосомные перестройки, моносомия и другие мутации, возникающие у одного из партнеров, способны вызвать значительные различия в фенотипе ОБ. Поэтому необходимо учитывать возможность соматических мутаций у ОБ в раннем эмбриогенезе.
Согласно обобщениям И. И. Канаева, изложенным в его превосходной монографии сущность близнецового метода в генетике сводится к следующим положениям:
1) пара ОБ имеет тождественную комбинацию, пара РБ - разные комбинации генотипов родителей;
2) для обоих партнеров одной пары ОБ внешняя среда может оказаться одинаковой, а для другой - разной. Если партнеры ОБ в течение жизни испытывают разное влияние, то это приведет к внутрипарному различию. Отсюда пары могут быть с внутрипарной одинаковой и внутрипарной разной средой.
Сравнение ОБ с одинаковой средой с ОБ с разной средой открывает возможность судить о роли влияния среды на внутрипарные различия близнецов в течение всей жизни. Сравнение ОБ с одинаковой средой и РБ с одинаковой средой позволяет выяснить роль наследственного фактора. Такого рода изучение проводят на большой выборке и обрабатывают статистически.
Исходя из разности генетического происхождения ОБ и РБ вытекает, что если по одним и тем же признакам нет различия у ОБ и есть таковые у РБ, то очевидно, что данные различия признаков у последних обусловлены наследственными факторами. Если же внутрипарные различия в тех же признаках встречаются у одного и другого типа близнецов, то очевидно, что они могут быть вызваны факторами среды. Из данных дискордантности у ОБ и РБ по ряду морфологических признаков, видно, что внутрипарное различие у РБ встречается во много раз чаще, чем у ОБ.

Представлены некоторые данные С. Рида относительно сравнительной частоты патологии у второго партнера в случае заболевания одного из близнецов.

В процентах показана частота конкордантности заболеваний у двух типов близнецов, из него видно, что если один партнер заболел одной из указанных болезней, то вероятность заболевания второго у ОБ значительно выше, чем у РБ. В. П. Эфроимсон, анализируя данные по частоте контордантных пар, совершенно правильно указывает, что высокая Наследственная предрасположенность ОБ к заболеваниям проявляется при наличии провоцирующего фактора; без него этот процент будет значительно ниже.
Близнецовый метод дает возможность с наибольшей точностью выяснить наследственную предрасположенность человека к ряду заболеваний и свойств. Другими методами очень трудно или почти невозможно исследовать многие инфекционные и опухолевые заболевания, воспаления кожи и различных органов, а также характеристики нормальной нервной деятельности человека.
При использовании близнецового метода приходится учитывать условия совместного и раздельного воспитания в жизни партнеров, социальные условия, в которых они находятся, и т. д. Тем не менее близнецовый метод позволяет наиболее точно определить, коэффициент наследуемости разных признаков, а также судить о гетерогенности популяции по изучаемым генам и вычленять роль среды в определении изменчивости изучаемых признаков.
Цитогенетический метод
Цитогенетическим методом в генетике человека обычно называют цитологический анализ кариотипа человека в норме и патологии.
Правильнее этот метод называть цитологическим, а не цитогенетическим, поскольку генетический анализ путем скрещивания у человека исключен, и носители хромосомных нарушений если выживают, то оказываются, как правило, бесплодными. Однако изредка в отношении некоторых хромосомных нарушений удается сочетать цитологический метод с генеалогическим и устанавливать связь фенотипического эффекта с определенным типом хромосомных изменений. В силу этих обстоятельств можно сохранить принятый в литературе термин «цитогенетический метод» в изучении генетики человека. В тех же случаях, где такого параллелизма исследовании не ведется, применение данного термина неправомочно.
Цитогенетическим методом исследуют различного рода гетероплоидию и хромосомные перестройки в соматических тканях человека, вызывающие различные фенотипические отклонения от нормы.
Чаще всего этот метод применяют на культуре ткани. Он позволяет учитывать крупные аномалии хромосом, возникающие как в половых, так и соматических клетках. Оказалось, что у человека, так же как и у животных, довольно часто возникают трисомики и моносомики по различным парам хромосом вследствие нерасхождения аутосом и половых хромосом в мейозе. Трисомия и моносомия по половым хромосомам у человека обнаруживаются на основе анализа полового хроматина.
В ходе относительно продолжительного индивидуального развития человека в клетках различных тканей накапливаются аномалии хромосом (хромосомные перестройки, а также изменение числа хромосом). Ткани организма представляют собой разнообразные популяции генетически различающихся клеток, в которых с возрастом концентрация клеток с патологическими ядрами возрастает. В этом случае цитогенетический метод позволяет изучать старение тканей на основе исследования структур клеток в возрастной динамике «популяции» соматических и генеративных тканей.
Поскольку частота возникновения хромосомных аномалий зависит от влияния на организм разнообразных мутагенов (ионизации, химических агентов - фармакологических препаратов, газового состава среды и др.), то цитогенетический метод позволяет устанавливать мутагенное действие факторов внешней среды на человека.
Применение цитогенетического метода особенно расширилось в связи с открытием причин ряда физических и психических заболеваний - так называемых хромосомных болезней.
Существует несколько заболеваний человека, например болезнь Клайнфельтера, Шерешевского-Тернера, Дауна и др., причины которых долго оставались неизвестными, пока цитологическими методами у таких больных не были обнаружены хромосомные аномалии.
Больные мужчины с синдромом Клайнфельтера характеризуются недоразвитием гонад, дегенерацией семенных канальцев, умственной отсталостью, непропорциональным ростом конечностей и т. д. У женщин встречается синдром Шерешевского-Тернера. Он проявляется в замедлении полового созревания, недоразвитии гонад, отсутствии менструаций, бесплодии, малом росте и в других Патологических признаках.
Оказалось, что оба эти синдрома у потомков являются следствием нерасхождения половых хромосом при образовании гамет родителей. Вследствие нерасхождения Х-хромосом у женского гомогаметного) пола в процессе мейоза могут возникать гаметы двумя Х-хромосомами, т. е. XX + 22 аутосомы, и без Х-хромосом, т. е. 0 + 22; у мужского (гетерогаметного) пола соответственно гаметы XY + 22 и 0 + 22. В случае оплодотворения таких яйцеклеток нормальными сперматозоидами (X + 22 или Y + 22) возможно образование следующих классов зигот: XXX + 44, 0Х + 44 и XXY + 44, 0Y + 44.
Из этого следует, что число хромосом у зигот разного происхождения может колебаться от 47 до 45, причем особи 0Y + 44, очевидно, не выживают, так как ни разу не были найдены. Хромосомный набор XXY + 44 присущ мужчине с синдромом Клайнфельтера (мужская интерсексуальность), хромосомные наборы Х0 + 44 и XXX + 44 имеют женщины с синдромом Шерешевского-Тернера.
При дальнейшем анализе больных с разными синдромами выяснилось, что вследствие нерасхождения половых хромосом могут возникать разного типа хромосомные аномалии, в частности полисомия. Встречаются, например, мужчины с такими наборами хромосом: XX Y, XXX Y, ХХХХ Y, а женщины - XXX, ХХХХ.
Особенность роли половых хромосом в детерминации пола у человека в случае их нерасхождения, в отличие от дрозофилы, проявилась в том, что набор хромосом XX Y всегда определяет мужской пол, а набор Х0 - женский. При этом увеличение числа Х-хромосом в сочетании с одной Y-хромосомой не изменяет определение мужского пола, а лишь усиливает синдром Клайнфельтера. Трисомия, или полисомия по Х-хромосоме, у женщин также часто вызывает заболевания, сходные с синдромом Шерешевского-Тернера.
Заболевания, вызванные нарушением нормального числа половых хромосом, диагностируются цитологическим методом - анализом полового хроматина. В тех случаях, когда в тканях мужчин имеется нормальный набор половых хромосом - XY, половой хроматин в клетках не обнаруживается. У нормальных женщин - XX - он обнаруживается в виде одного тельца. При полисомии по Х-хромосомам у женщин и мужчин количество телец полового хроматина всегда на единицу меньше числа Х-хромосом, т. е. n x = n·Х - 1. Так, в клетках мужчин с синдромом Клайнфельтера при наборе XX Y имеется одно тельце полового хроматина, при наборе XXXY - два, при наборе XXXXY - три; у женщин с синдромом Шерешевского-Тернера соответственно: Х0 - нет тельца, XXX - два тельца, ХХХХ - три тельца полового хроматина, и т. д. Предполагается, что в каждой такой зиготе генетически активна лишь одна из Х-хромосом. Остальные хромосомы переходят в гетеропикнотическое состояние в виде полового хроматина.
Причины этой закономерности не выяснены, однако предполагается, что она связана с нивелированием действия генов половых хромосом у гетеро- и гомогаметного пола.
Как мы знаем, нерасхождение хромосом может происходить не только в мейозе, но и в соматических клетках в течение всего эмбриогенеза животного начиная с первых дроблений яйца. В силу последнего среди людей при нарушении расхождения половых хромосом могут появиться больные мозаики-женщины и мозаики-мужчины. Так, например, описаны мозаики следующих типов: двойные: Х0/XX, Х0/XXX и X0/XY, X0/XYY, тройные: Х0/ХХ/ХХХ, XX/X0/XY, а также четверные мозаики, когда соматические клетки одного человека содержат четыре разных набора хромосом.
Кроме рассмотренного типа болезней, вызванных изменением числа половых хромосом в зиготе, хромосомные болезни могут быть вызваны нерасхождением аутосом и разного рода хромосомными перестройками (транслокациями, делециями). Так, например, у детей с врожденной идиотией - болезнью Дауна, сопровождающейся малым ростом, широким круглым лицом, близко расположенным узкими глазными щелями и полуоткрытым ртом, была обнаружена трисомия по 21 хромосоме. Установлено, что частота встречаемости болезни Дауна у новорожденных зависит от возраста матерей.
С врожденными хромосомными аномалиями связывают весьма разнообразные болезни. Поэтому цитогенетический метод приобретает важное значение в этиологии болезней человека.
Популяционный метод
Популяционный метод позволяет изучать распространение отдельных генов или хромосомных аномалий в человеческих популяциях.
Популяционный метод основывается на математических методах. Для анализа генетической структуры популяции необходимо обследовать большую по размеру выборку, которая должна быть репрезентативной - объективно отражать всю генеральную совокупность, т. е. всю популяцию в целом. В обследуемой выборке устанавливают распределение лиц по соответствующим четко очерченным фенотипическим классам, различия между которыми наследственно обусловлены. Затем, исходя из найденных фенотипических частот, определяют генные частоты.
На основе знания генных частот представляется возможность дать описание анализируемой популяции в соответствии с формулой Гарди-Вайнберга и заранее предсказать вероятный характер расщепления в потомстве лиц, относящихся к тем или иным фенотипическим классам. Исследование генных частот имеет важное значение для оценки последствий родственных браков, а также для выяснения генетической истории человеческой популяции в целом.
Частота распространения в популяциях разных аномалий оказывается различной; при этом подавляющее количество соответствующих рецессивных аллелей представлено в гетерозиготном состоянии.

Так, примерно каждый сотый житель Европы гетерозиготен по гену амавротической идиотии (болезнь Шпильмайера-Фогта), тогда как заболевают этой болезнью в юношеском возрасте из 1 млн. только 25 человек, являющихся гомозиготными. Альбиносы в европейских странах встречаются с частотой 1 на 20 000, хотя гетерозиготное состояние этой аллели присуще каждому семидесятому жителю.
Несколько иначе дело обстоит в случае аномалий, наследующихся сцеплено с полом, примером чего может служить дальтонизм - цветная слепота, которая контролируется, по-видимому, рядом аллелей, распределенных по двум тесно сцепленным локусам в Х-хромосоме. Среди мужского населения частота дальтоников (q) соответствует суммарной частоте рецессивных аллелей и составляла, например, в Москве в 30-х годах, по данным Р. И. Серебровской, 7%, в то же время среди женского населения той же популяции цветная слепота была только у 0,5% (q 2), но в гетерозиготном состоянии примерно 13% женщин несут аллели, обусловливающие дальтонизм.
Как мы уже говорили выше, рассматривая генеалогический метод, вероятность появления в потомстве рецессивных гомозигот может быть различной при вступлении в брак лиц, имеющих разную степень родства. Так, у супругов, являющихся по отношению друг к другу двоюродными братьями и сестрами, вероятность рождения детей, гомозиготных по рецессивной аллели, распространенной в популяции с частотой q, составит уже не q 2 , а большую величину, а именно q/16 (1 +15q).
Это связано с тем, что если один из общих предков таких супругов - бабушка или дедушка - нес в гетерозиготе рецессивную аллель, то с вероятностью 1/16 данная аллель передастся обоим двоюродным сибсам.
Вредные последствия родственных браков особенно наглядно проявляются в изолированных популяциях ограниченного размера, так называемых изолятах . Под изолятом понимают группу особей популяции, которые вступают в брак большей частью с особями своей группы и поэтому характеризуются значительным коэффициентом кровного родства. Такими изолятами могут быть отдельные изолированные селения, общины и т. д. Внутри изолята более вероятны родственные браки (инбридинг), и больше шансов на то, что супруги будут нести одинаковые мутантные гены, следствием чего является увеличение вероятности проявления рецессивных аллелей в гомозиготном состоянии. Разные изоляты несут различные концентрации сходных или разных генов.
На Марианских островах и острове Гуам смертность среди местного населения от бокового амиотрофического склероза (связанного с поражением клеток передних рогов спинного мозга) в 100 с лишним раз превышает смертность от этой болезни в других странах. В Южной Панаме в провинции Сан-Блаз весьма заметную часть племени кариба куна составляют альбиносы, которые появляются здесь в каждом поколении. В одном селении на р. Роне в Швейцарии среди 2200 жителей имеется более 50 глухонемых, и еще у 200 обнаруживаются некоторые дефекты слуха. По всей вероятности, во всех подобных случаях резкого увеличения концентрации отдельных аллелей известную роль играет генетический дрифт, неравномерное размножение в прошлом отдельных семей, родов, а также снижение миграции.
По мере роста цивилизации и развития производительных сил общества количество изолятов уменьшается, и их значение для популяции в целом падает. Однако они все еще имеют место.
Знание генных частот, как уже говорилось позволяет предсказывать характер расщепления в потомстве отдельных фенотипических классов родительских особей.
Исходя из формулы Гарди-Вайнберга, можно показать, что при моногенном наследовании расщепление по фенотипу в потомстве доминантных матерей должно осуществляться в соотношении p(1 + pq) доминантов к р рецессивов, или (l+pq):q 2 ; в потомстве рецессивных матерей расщепление по фенотипу должно быть pq 2: q 3 , или p: q.
Приведем пример. В одном исследовании при изучении резус-фактора частота рецессивной аллели rh в популяции составила 0,4, а частота доминантной аллели Rh - 0,6. Отсюда следовало ожидать, что в потомстве резус-положительных матерей частота резус — положительных детей (Rh +) примерно в 7,8 раза будет превышать частоту резус-отрицательных детей (Rh —); в потомстве резус-отрицательных матерей соответствующее превышение будет в 1,5 раза.
Действительные соотношения в обследованной выборке составили:
- в первом случае 1475 Rh + : 182 Rh — , или 8,1: 1,
- во втором случае 204 Rh + : 129 Rh — , или 1,6: 1.
Таким образом, наблюдаемые результаты при расщеплении весьма хорошо соответствуют теоретически ожидаемым результатам, предсказанным на основе анализа генных частот.
Популяционный анализ полиморфизма по группам крови интересен тем, что он помогает понять динамику генетической структуры различных популяций и способствует выявлению связей между ними.
Разные популяции существенно различаются по своей генетической структуре, в частности по группам крови. При этом удается проследить некоторые вполне четкие закономерности. Если концентрация аллели I B наибольшая в районе Индии и Китая, то к востоку и западу от этого района происходит постепенное падение ее вплоть до нуля среди коренных обитателей Америки и Австралии. В то же время у американских индейцев (и аборигенов Австралии и Полинезии) максимума достигает концентрация аллели I 0 . Аллель I А редка у коренного населения Америки, а также в Индии, Аравии, тропической Африке, в Западной Европе.
Для объяснения этих различий в генетической структуре популяций недавно была предложена гипотеза, согласно которой решающим фактором отбора в отношении групп крови системы АВ0 явились эпидемии чумы и оспы. Возбудитель чумы Pasteuvella pest is, обладая свойством антигена 0, оказывается наиболее губительным для людей с группой крови 0, поскольку такие лица не способны вырабатывать достаточное количество антител в случае инфекции. По аналогичной причине вирус оспы наиболее опасен для людей с группой крови А. Там, где свирепствовала чума (Индия, Монголия, Китай, Египет), шла интенсивная элиминация аллели I 0 , а там, где особенно свирепствовала оспа (Америка, Индия, Аравия, тропическая Африка), в первую очередь элиминировалась аллель 1 А. В районах Азии, где чума и оспа были эндемичны, наибольшую частоту получила аллель 1 в.
В главе 5 мы рассмотрели моногенное наследование серповидноклеточной анемии, обусловленное расщеплением по аллелям гена S. Высокая концентрация аллели S в поясе эндемичной малярии (Африка, Средиземноморье) оказалась связанной с повышенной устойчивостью к малярии гетерозигот (Ss) и с возникновением. в результате этого системы сбалансированного наследственного полиморфизма.
Таким образом, в обоих приведенных примерах анализа полиморфизма по группам крови и серповидно-клеточной анемии мы видим, как применение популяционного метода позволяет вскрывать генетическую структуру человеческих популяций.
Онтогенетический метод
Онтогенетический метод позволяет устанавливать по фенотипу носительство рецессивных аллелей в гетерозиготном состоянии и хромосомных перестроек.
Генетической основой проявления рецессивных генов в гетерозиготном состоянии является, по-видимому, неполный блок в цепи синтеза того или иного метаболита, вызванного действием доминантной аллели данного гена.
Известно, что некоторые наследственные болезни проявляются не только у лиц, гомозиготных по аллелям, вызывающим заболевание, но в стертой форме и у гетерозигот. Поэтому в настоящее время усиленно разрабатываются методы определения гетерозиготного носительства в онтогенезе. Так, гетерозиготный носитель фенилкетонурии (повышенное содержание фенилаланина в крови определяется дополнительным введением фенилаланина и последующим определением уровня его (или тирозина) в плазме крови. Наличие гетерозиготности по данной аллели устанавливается по повышенному содержанию фенилаланина. В норме (т. е. у гомозигот по доминантной аллели) уровень фенилаланина не изменяется. В норме в крови присутствует фермент каталаза, необходимый для углеводного обмена, но встречается ген, который в гомозиготном состоянии вызывает отсутствие каталазы. У гомозиготных носителей этого гена наблюдается болезнь акаталаземия - расстройство углеводного обмена. Гетерозиготы занимают промежуточное положение по активности каталазы без большого захождения между доминантными и рецессивными гомозиготами.
По активности каталазы можно точно определить гетерозиготных и гомозиготных носителей аллели акаталаземии среди близких родственников и родителей.
Гетерозиготное носительство аллели, определяющей мышечную дистрофию типа Дюшена, тестируется по активности криатинфосфокиназы. Теперь разработаны, подобные тесты для 40 наследственных болезней, определяемых рецессивными аллелями.
В настоящее время онтогенетический метод обогатился за счет биохимических, иммунологических и молекулярных приемов исследования, описанию которых посвящен ряд специальных руководств.
Важность онтогенетического метода очевидна для установления носительства рецессивного гена в гетерозиготном состоянии у родственников семьи, в которой появляется наследственно больной ребенок. Диагностика в онтогенезе важна для расчета вероятности появления наследственно больных потомков при родственных и смешанных браках. По мере упрощения тестирования гетерозиготного носительства этот метод должен будет внедряться в целях консультации супружеских пар относительно возможности появления заболевания у их детей, а также для изучения распространения мутаций в популяциях.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
В основе этого метода лежит составление и анализ родословных. Этот метод широко применяют с древних времен и до наших дней в коневодстве, селекции ценных линий крупного рогатого скота и свиней, при получении чистопородных собак, а также при выведении новых пород пушных животных. Родословные человека составлялись на протяжении многих столетий в отношении царствующих семейств в Европе и Азии.
Как метод изучения генетики человека генеалогический метод стали применять только с началаXX столетия, когда выяснилось, что анализ родословных, в которых прослеживается передача из поколения в поколение какого-то признака (заболевания), может заменить собой фактически неприменимый в отношении человека гибридологический метод.
При составлении родословных исходным является человек - пробанд, родословную которого изучают. Обычно это или больной, или носитель определенного признака, наследование которого необходимо изучить. При составлении родословных таблиц используют условные обозначения, предложенные Г. Юстом в 1931 г. (рис. 6.24). Поколения обозначают римскими цифрами, индивидов в данном поколении - арабскими.
Рис. 6.24. Условные обозначения при составлении родословных (по Г. Юсту)
С помощью генеалогического метода может быть установлена наследственная обусловленность изучаемого признака, а также тип его наследования (аутосомно-доминантный, аутосомно-рецессивный, X-сцепленный доминантный или рецессивный, Y-сцепленный). При анализе родословных по нескольким признакам может быть выявлен сцепленный характер их наследования, что используют при составлении хромосомных карт. Этот метод позволяет изучать интенсивность мутационного процесса, оценить экспрессивность и пенетрантность аллеля. Он широко используется в медико-генетическом консультировании для прогнозирования потомства. Однако необходимо отметить, что генеалогический анализ существенно осложняется при малодетности семей.
Родословные при аутосомно-доминантном наследовании. Для аутосомного типа наследования в целом характерна равная вероятность встречаемости данного признака как у мужчин, так и у женщин. Это обусловлено одинаковой двойной дозой генов, расположенных в аутосомах у всех представителей вида и получаемых от обоих родителей, и зависимостью развивающегося признака от характера взаимодействия аллельных генов.
При доминировании признака в потомстве родительской пары, где хотя бы один родитель является его носителем, он проявляется с большей или меньшей вероятностью в зависимости от генетической конституции родителей (рис. 6.25).

Рис. 6.25. Вероятность появления потомков с доминантным признаком от различных супружеских пар (/-III )
Если анализируется признак, не влияющий на жизнеспособность организма, то носители доминантного признака могут быть как гомо-, так и гетерозиготами. В случае доминантного наследования какого-то патологического признака (заболевания) гомозиготы, как правило, нежизнеспособны, а носители этого признака - гетерозиготы.
Таким образом, при аутосомно-доминантном наследовании признак может встречаться в равной мере у мужчин и у женщин и прослеживается при достаточном по численности потомстве в каждом поколении по вертикали. Анализируя родословные, необходимо помнить о возможности неполного пенетрирования доминантного аллеля, обусловленной взаимодействием генов или факторами среды. Показатель пенетрантности может быть вычислен как отношение фактического числа носителей признака к числу ожидаемых носителей этого признака в данной семье. Необходимо также помнить, что некоторые заболевания проявляются не сразу с момента рождения ребенка. Многие болезни, наследуемые по доминантному типу, развиваются лишь в определенном возрасте. Так, хорея Гентингтона клинически проявляется к 35-40 годам, поздно проявляется и поликистоз почек. Поэтому при прогнозировании подобных заболеваний в расчет не принимаются братья и сестры, не достигшие критического возраста.
Первое описание родословной с аутосомно-доминантным типом наследования аномалии у человека было дано в 1905 г. В ней прослеживается передача в ряду поколений брахидактилии (короткопалости). На рис. 6.26 приведена родословная с этой аномалией. На рис. 6.27 изображена родословная с ретинобластомой в случае неполной пенетрантности.
Родословные при аутосомно-рецессивном наследовании. Рецессивные признаки проявляются фенотипически лишь у гомозигот по рецессивным аллелям. Эти признаки, как правило, обнаруживаются у потомков фенотипически нормальных родителей - носителей рецессивных аллелей. Вероятность появления рецессивного потомства в этом случае равна 25%. Если один из родителей имеет рецессивный признак, то вероятность проявления его в потомстве будет зависеть от генотипа другого родителя. У рецессивных родителей все потомство унаследует соответствующий рецессивный признак (рис. 6.28).

Рис. 6.26. Родословная (А ) при аутосомно-доминантном типе наследования (брахидактилия - Б )
Для родословных при аутосомно-рецессивном типе наследования характерно, что признак проявляется далеко не в каждом поколении. Чаще всего рецессивное потомство появляется у родителей с доминантным признаком, причем вероятность появления такого потомства возрастает в близкородственных браках, где оба родителя могут являться носителями одного и того же рецессивного аллеля, полученного от общего предка. Примером аутосомно-рецессивного наследования является родословная семьи с псевдогипертрофической прогрессивной миопатией, в которой часты близкородственные браки (рис. 6.29). Обращает внимание распространение заболевания в последнем поколении по горизонтали.

Рис. 6.27. Родословная с ретинобластомой в случае неполной пенетрантности

Рис. 6.28. Вероятность появления потомков с рецессивным признаком
от различных супружеских пар (I-IV )
Родословные при доминантном Х-сцепленном наследовании признака. Гены, расположенные в Х-хромосоме и не имеющие аллелей в Y-хромосоме, представлены в генотипах мужчин и женщин в разных дозах. Женщина получает две свои Х-хромосомы и соответствующие гены как от отца, так и от матери, а мужчина наследует свою единственную Х-хромосому только от матери. Развитие соответствующего признака у мужчин определяется единственным аллелем, присутствующим в его генотипе, а у женщин он является результатом взаимодействия двух аллельных генов. В связи с этим признаки, наследуемые по Х-сцепленному типу, встречаются в популяции с разной вероятностью у мужского и женского пола.
При доминантном Х-сцепленном наследовании признак чаще встречается у женщин в связи с большей возможностью получения ими соответствующего аллеля либо от отца, либо от матери. Мужчины могут наследовать этот признак только от матери. Женщины с доминантным признаком передают его в равной степени дочерям и сыновьям, а мужчины - только дочерям. Сыновья никогда не наследуют от отцов доминантного Х-сцепленного признака.

Рис. 6.29. Родословная при аутосомно-рецессивном типе наследования (псевдогипертрофическая прогрессирующая миопатия)
Примером такого типа наследования служит описанная в 1925 г. родословная с фолликулярным кератозом - кожным заболеванием, сопровождающимся потерей ресниц, бровей, волос на голове (рис. 6.30). Характерным является более тяжелое течение заболевания у гемизиготных мужчин, чем у женщин, которые чаще всего являются гетерозиготами.
При некоторых заболеваниях наблюдается гибель мужчин-гемизигот на ранних стадиях онтогенеза. Тогда в родословных среди пораженных должны быть только женщины, в потомстве которых отношение пораженных дочерей, здоровых дочерей и здоровых сыновей равно 1:1:1. Мужские доминантные гемизиготы, не погибающие на очень ранних стадиях развития, обнаруживаются в самопроизвольных абортах или среди мертворожденных. Такими особенностями наследования у человека характеризуется пигментный дерматоз.
Родословные при рецессивном Х-сцепленном наследовании признаков. Характерной особенностью родословных при данном типе наследования является преимущественное проявление признака у гемизиготных мужчин, которые наследуют его от матерей с доминантным фенотипом, являющихся носительницами рецессивного аллеля. Как правило, признак наследуется мужчинами через поколение от деда по материнской линии к внуку. У женщин он проявляется лишь в гомозиготном состоянии, вероятность чего возрастает при близкородственных браках.
Наиболее известным примером рецессивного Х-сцепленного наследования является гемофилия. Наследование гемофилии типа А представлено в родословной потомков английской королевы Виктории (рис. 6.31).

Рис. 6.30. Родословная при Х-сцепленном доминантном типе наследования (фолликулярный кератоз)

Рис. 6.31. Родословная при Х-сцепленном рецессивном типе наследования (гемофилия типа А)
Другим примером наследования по данному типу является дальтонизм - определенная форма нарушения цветоощущения.
Родословные при Y-сцепленном наследовании. Наличие Y-хромосомы только у представителей мужского пола объясняет особенности Y-сцепленного, или голандриче-ского, наследования признака, который обнаруживается лишь у мужчин и передается по мужской линии из поколения в поколение от отца к сыну.

Рис. 6.32. Родословная при Y-сцепленном (голандрическом) типе наследования
Одним из признаков, Y-сцепленное наследование которого у человека все еще обсуждается, является гипертрихоз ушной раковины, или наличие волос на внешнем крае ушной раковины. Предполагают, что в коротком плече Y-хромосомы кроме этого гена находятся гены, определяющие мужской пол. В 1955 г. у мыши описан определяемый Y-хромосомой трансплантационный антиген, названный HY. Возможно, он является одним из факторов половой дифференцировки мужских гонад, клетки которых имеют рецепторы, связывающие этот антиген. Связанный с рецептором антиген активизирует развитие гонады по мужскому типу (см. разд. 3.6.5.2; 6.1.2). Этот антиген в процессе эволюции остался почти неизменным и встречается в организме многих видов животных, в том числе и человека. Таким образом, наследование способности к развитию гонад по мужскому типу определяется голандрическим геном, расположенным в Y-хромосоме (рис. 6.32).
Близнецовый метод
Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов. Он предложен в 1875 г. Гальтоном первоначально для оценки роли наследственности и среды в развитии психических свойств человека. В настоящее время этот метод широко применяют в изучении наследственности и изменчивости у человека для определения соотносительной роли наследственности и среды в формировании различных признаков, как нормальных, так и патологических. Он позволяет выявить наследственный характер признака, определить пенетрантность аллеля, оценить эффективность действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания).
Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны, так как имеют 100% общих генов. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордантных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбрионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам между близнецами наблюдается дискордантность, т.е. различия. Напротив, сохранение сходства между близнецами, несмотря на различия условий их существования, свидетельствует о наследственной обусловленности признака.
Сопоставление парной конкордантности по данному признаку у генетически идентичных монозиготных и дизиготных близнецов, которые имеют в среднем около 50% общих генов, дает возможность более объективно судить о роли генотипа в формировании признака. Высокая конкордантность в парах монозиготных близнецов и существенно более низкая конкордантность в парах дизиготных близнецов свидетельствуют о значении наследственных различий в этих парах для определения признака. Сходство показателя конкордантности у моно- и дизиготных близнецов свидетельствует о незначительной роли генетических различий и определяющей роли среды в формировании признака или развития заболевания. Достоверно различающиеся, но достаточно низкие показатели конкордантности в обеих группах близнецов дают возможность судить о наследственной предрасположенности к формированию признака, развивающегося под действием факторов среды.
Установление соотносительной роли наследственности и среды в развитии различных патологических состояний позволяет врачу правильно оценить ситуацию и проводить профилактические мероприятия при наследственной предрасположенности к заболеванию или осуществлять вспомогательную терапию при его наследственной обусловленности.
Трудности близнецового метода связаны, во-первых, с относительно низкой частотой рождения близнецов в популяции (1:86-1:88), что осложняет подбор достаточного количества пар с данным признаком; во-вторых, с идентификацией монозиготности близнецов, что имеет большое значение для получения достоверных выводов.
Для идентификации монозиготности близнецов применяют ряд методов. 1. Полисимптомный метод сравнения близнецов по многим морфологическим признакам (пигментации глаз, волос, кожи, форме волос и особенностям волосяного покрова на голове и теле, форме ушей, носа, губ, ногтей, тела, пальцевым узорам). 2. Методы, основанные на иммунологической идентичности близнецов по эритроцитарным антигенам (системы АВО, MN, резусу), по сывороточным белкам (γ-глобулину). 3. Наиболее достоверный критерий монозиготности предоставляет трансплантационный тест с применением перекрестной пересадки кожи близнецов.
Несмотря на трудоемкость близнецового метода и возможность ошибок при определении монозиготности близнецов, высокая объективность выводов делает его одним из широко применяемых методов генетических исследований у человека.
Разделы: Биология
Учебно-исследовательская работа студентов (УИРС) в Черемховском медицинском техникуме в период профессиональной подготовки является одной из основных форм самостоятельной работы студентов
УИРС – является одним из активных методов обучения деятельностного характера, что соответствует новым требованиям ФГОС. Студенту в процессе профессиональной подготовки самостоятельно приходится из большого числа научно – методической и профильной литературы находить отдельные теоретические выкладки, а также самостоятельно проводить инструментальные и лабораторные исследования с последующим анализом полученных результатов.
При проведении УИРС у студентов формируются определенные общекультурные и профессиональные компетенции, через развитие интеллектуальных и профессиональных умений (работать с литературой разного характера, выделять главное, уметь анализировать, планировать свою деятельность, выдвигать предположение, проводить исследование, анализировать результаты, делать выводы и.т.д).
УИРС – это собственный творческий труд студента с итоговыми выводами и суждениями по работе, где студенты выражают свой потенциал будущего исследователя, проявляя интерес к исследовательской работе и понимание её необходимости.
Представляемая работа выполнена в соответствии с требованиями к УИРС.
Цель исследования
Практическая значимость: Обучение навыкам составления и анализа родословной. Разработка памятки по составлению и анализу родословных. Просвещение студентов по вопросам генеалогии, развитие интереса к более глубокому изучению проблемы.
Генеалогический метод как универсальный метод изучения наследственности человека
Ковальчук Елена
студентка 2 курса, специальность «Сестринское дело»
Областное государственное бюджетное образовательное учреждение
среднего профессионального образования
«Черемховский медицинский техникум»
научный руководитель – Склярова Светлана Владимировна
Введение
В настоящее время, по данным Всемирной организации здравоохранения известно около 10 тысяч наследственных заболеваний, которые приобретают все больший удельный вес в общей патологии человека. Основной причиной наследственных болезней считают вредные мутации генов. Изучением наследственных заболеваний человека занимается медицинская генетика. Для диагностики наследственной патологии в медицинской генетике применяется генеалогический метод. Этот метод является доступным и информационным, он даёт возможность установить наследуемый характер заболевания, тип передачи дефектного гена, проследить возможный риск проявления его у близких родственников.
Выбор темы обусловлен интересом к изучению родословной моей семьи, так как в нашем роду отмечаются часто повторяющиеся заболевание, возникла необходимость изучения его наследственной природы.
Цель исследования : использование генеалогического метода с целью выявления наследственных заболеваний в роду.
Объект исследования: родословная семьи Ковальчук Елены Игоревны по линии матери.
Задачи исследования:
- Проанализировать научную основу генеалогического метода.
- Путем практического применения метода составить родословную семьи.
- Сделать анализ родословной, выявить характер и тип наследуемых признаков.
- Разработать памятку по составлению и анализу родословных.
М етоды исследования: изучение и анализ общей и специальной литературы, наблюдение, метод интервью, качественный анализ родословной.
Практическая значимость: Обучение навыкам составления и анализа родословной. Разработка памятки по составлению и анализу родословных. Просвещение студентов по вопросам генеалогии.
Глава 1. Генеалогический метод изучения наследственности человека
Таким образом, генеалогический метод, широко применяется при решении теоретических и прикладных проблем: установление наследственного характера признака; определение типа наследования заболевания. Определить прогноз заболевания и рассчитать риск для потомства.
В генеалогическом методе, можно выделить 2 этапа: 1этап – составление родословных; 2 этап – использования генеалогических данных для генетического анализа.
Глава 2. Составление и анализ родословной
Таким образом, составление родословной с учетом основных правил и требований позволит успешно провести качественный анализ родословной, который в свою очередь даст наиболее полную информацию о характере и типе наследуемого признака, а также определит вероятность передачи признака последующим поколениям.
Глава 3. Критерии типов наследования
Таким образом, изучив критерии типов и особенности наследования признаков становиться возможным, более точно установить характер наследования признаков в изучаемой родословной и предположить вероятность проявление гена в последующих поколениях
Глава 4. Родословная и ее анализ
4.1 Составление родословной
С целью выявления наличия в роду наследуемых болезней составлена родословная, с учетом основных требований (Приложение ).
Определена «легенда родословной», включающая в себя: краткую запись с точной характеристикой членов семьи и его родства с пробандом, сведения о состоянии здоровья членов родословной, информацию о характере наследования заболевания и особенностях его проявления, о начале и характере течения заболевания, о возрасте. Информация получена методом опроса родственников, прежде всего родителей, а также бабушек и дедушек. Собранная информация позволила провести анализ родословной, а именно установить наследуемый ли признак, а также понять характер наследования данного заболевания.
4.2 Анализ родословной
С целью установления наследственных закономерностей проведен генетический анализ родословной, который показал:
В первом, третьем и четвертом поколениях по вертикальному направлению отмечаются по одному случаю заболевания тонзиллитами – это говорит о наследственном характере признака, так как это повторяющиеся случаи заболевания. Тонзиллит не является наследственным заболеванием, поэтому определяется наследственная предрасположенность к данному заболеванию, в основе которого лежит снижение иммунного ответа на возбудитель заболевания.
Установлен аутосомно-доминантный тип наследования признака, так как в первом, третьем и четвертом поколениях отмечается по одному случаю заболевания тонзиллитом у женщин, то есть наблюдается прямая передача признака от одного из больных родителей к детям, в данном случае от матери к ребенку (дочерям) – это свойственно для данного типа наследования признака.
Данный тип наследования подтверждает тот факт, что во втором поколении заболевание не проявилось, это говорит о неполной пенетрантности потомков больного человека, то есть человек, будучи внешне здоровым, но он передает своим детям гены, отвечающие за это заболевание, либо предрасположенность к нему, как в нашем случае.
Для данного типа наследования характерно нарастание тяжести патологических нарушений в последующих поколениях, которых поддаются коррекции путем профилактических мероприятий.
Таким образом, результаты анализа родословной позволили установить:
- Характер наследуемого признака – наследственная предрасположенность к снижению иммунного ответа на возбудитель тонзиллита;
- Определить тип наследуемого признака как аутосомно-доминантный.
- Предположить, что последующие поколения от пробанда могут наследовать данный признак.
- Для избегания нарастания тяжести патологических нарушений в последующих поколениях необходимо проводить профилактические мероприятия.
Заключение
Данное исследование было направлено на применение генеалогического метода с целью выявления наследственных заболеваний в роду.
По данной проблеме изучена специальная литература, содержание которой отражает научную основу генеалогического метода. Теоретическое изучение вопроса указывает на актуальность изучения генетики человека в связи с учащением проявления наследственных заболеваний, в том числе на своевременную диагностику наследственных заболеваниях.
Важную роль в диагностики данной категории заболеваний отводят генеалогическому методу. Данный метод характеризует высокая эффективность, так как является наиболее информационным, а также доступным для любого человека, интересующимся историей развития своей семьи или рода, в том числе наличием в роду наследственных заболеваний.
В процессе применения генеалогического метода на практике составлена родословная и сделан ее качественный анализ. Результаты анализа показали:
- Наличие в роду наследственной предрасположенности к тонзиллитам, в основе которого лежит снижение иммунного ответа на возбудитель.
- Предрасположенность к заболеванию передаётся по женской вертикальной линии.
- Наследование признака относится к аутосомно-доминантному типу наследования.
- При данном типе наследования признака характерно нарастание тяжести патологических нарушений в последующих поколениях.
Таким образом, анализ родословной позволяет понять ее наследственную природу, то есть удалось установить характер и тип наследуемого признака.
Генеалогический метод подтверждает свою универсальность, так как позволил определить характер и тип наследования признака, предположить риск для будущих поколений. Остается наиболее доступным и информационным методом в диагностике генетических заболеваний.
В ходе работы, по результатам исследования установлено, что проявление генетических заболеваний, а также снижение нарастания тяжести патологических нарушений в последующих поколениях можно избежать, осуществляя профилактические мероприятия.
Соблюдение рекомендуемых профилактических мероприятий, обеспечивающие ведение здорового образа жизни, позволят предупредить частые обострения заболевания, снизить риск развития нарастания тяжести патологических нарушений в последующих поколениях и соответственно снизит вероятность передачи пробандом мутантного гена последующим поколениям.
Таким образом, здоровый образ жизни является залогом предупреждения проявления у человека не только ненаследственных, но и генетических заболеваний.